


GW bodies, microRNA, and innate immune response
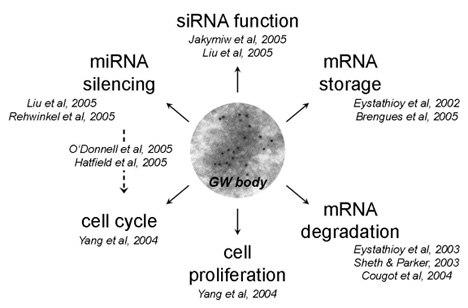
GW bodies (GWBs) are cytoplasmic foci initially identified through the use of an autoimmune serum targeting the marker protein GW182 (Mol Biol Cell. 2002; 13:1338-51). GWBs were first considered as both storage centers for a specific subset of mRNAs and degradation sites for mRNAs (RNA. 2003; 9:1171-3). They are known to vary in size and number throughout the cell cycle and are largest in size and most abundant in number during the late S and G2 phases (Proc Natl Acad Sci U S A. 1993; 90:1947-51). Our earlier studies linked RNA interference to GWBs, in that disruption or disassembly of GWBs was demonstrated to impair siRNA and miRNA silencing activity (Nat Cell Biol. 2005; 7:1267-74). As miRNAs are implicated in the regulation of cell cycle progression and cell proliferation, it is likely that GWBs, the critical intracellular structures for miRNA function, may very well be linked to this cellular process.
A book titled “Ten Years of Progress in GW/P Body Research” was published in 2012 and edited by Drs. Edward Chan and Marvin J. Fritzler.
Dominant microRNA in innate immune response regulating endotoxin tolerance
Our 2009 JBC article (J Biol Chem. 2009; 284:34590-9) was the first to define the critical role of miR-146a in endotoxin tolerance. We have followed up with several studies to document that miR-146a has a dominant role in IL-1β-induced tolerance and cross-tolerance to toll-like receptor (TLR) ligands (J Immunol. 2011; 186:1723-34; J Innate Immun. 2015; 7:428-40). TLR and IL-1R signaling activates the MyD88-dependent pathway with the formation of the myddosome, involving the helical assembly of the MyD88-IRAK4-IRAK2/IRAK1 complex. TLR/IL-1R signaling leads to activation of NF-κB, which is known to activate miR-146a production. TLR2 and TLR5 ligands also activate transcription factor CREB and miR-132/-212 production. miR-132/-212 targets IRAK4 (J Immunol. 2013; 190:1250-63), whereas miR-146a targets IRAK2/1 and TRAF6. IRAK4 and IRAK2/1 are critical components of the myddosome, which is in turn critical for activation of the pathway. Thus, these miRNA operate as a negative regulatory feedback mechanism to prevent the destructive consequences of uncontrolled cytokine production during the IL-1β/TLR signaling cascade. One of our most highly cited works is the early identification of elevated miR-146a in peripheral blood mononuclear cells of patients with rheumatoid arthritis and the high level of this microRNA correlated with disease activity (Arthritis Res Ther. 2008; 10:R101). We also showed that GW bodies are formed and can be used as markers for microRNA-mediated regulation of innate immune signaling in THP-1 cells. (Immunol Cell Biol. 2010; 88:205-12).
Publications
Vracar TC, Zuo J, Park J, Azer D, Mikhael C, Holliday SA, Holsey D, Han G, VonMoss L, Neubert JK, Rody WJ Jr, Chan EKL, and Holliday LS. Enoxacin and bis-enoxacin stimulate 4T1 murine breast cancer cells to release extracellular vesicles that inhibit osteoclastogenesis. Sci Rep. 2018; 8:16182.
Nahid, M.A., Satoh, M., and Chan, E.K. Interleukin 1β-Responsive MicroRNA-146a Is Critical for the Cytokine-Induced Tolerance and Cross-Tolerance to Toll-Like Receptor Ligands. J Innate Immun. 2015; 7:428-40.
Fredenburg, K.M. and Chan, E.K. Our journey from the study of human autoantibodies to the microRNA world. Front Immunol. 2015; 6:110. Review.
Nahid, M.A., Yao, B., Dominguez-Gutierrez, P.R., Kesavalu, L., Satoh, M., and Chan, E.K. Regulation of TLR2-mediated tolerance and cross-tolerance through IRAK4 modulation by miR-132 and miR-212. J Immunol. 2013; 190:1250-63.
Yao, B., La, L.B., Chen, Y.C., Chang, L.J., and Chan, E.K. Defining a new role of GW182 in maintaining miRNA stability. EMBO Rep. 2012; 13:1102-8.
Yao, B., Li, S., Lian, S.L., Fritzler, M.J., and Chan, E.K. Mapping of Ago2-GW182 functional interactions. Methods Mol. Biol. 2011; 725:45-62.
Nahid, M.A., Satoh, M., and Chan, E.K. Mechanistic role of miR-146a in endotoxin-induced differential cross-regulation of TLR signaling. J Immunol. 2011; 186:1723-34.
Yao, B., Li. S., Jung, H.M., Lian, S.L., Abadal, G.X., Han, F., Fritzler, M.J., and Chan, E.K. Divergent GW182 functional domains in the regulation of translational silencing. Nucleic Acids Res. 2011; 39:2534-47.
Ceribelli, A., Tincani, A., Cavazzana, I., Franceschini, F., Cattaneo, R., Pauley, B.A., Chan, J.Y., Chan, E.K., and Satoh, M. Anti-argonaute2 (Ago2/Su) and -Ro antibodies identified by immunoprecipitation in primary anti-phospholipid syndrome (PAPS). Autoimmunity. 2011; 44:90-7.
Nahid, M.A., Rivera, M., Lucas, A., Chan, E.K., and Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances miR-146a in ApoE-/- mice during experimental periodontal disease. Infection Immun. 2011; 79:1597-605.
Vázquez-Del Mercado, M., Sánchez Orozco, L.V., Pauley, B.A., Chan, J.Y., Chan, E.K., Panduro, A., Maldonado González, M., Jimenez-Luevanos, M.A., Martin Márquez, B.T., Palafox-Sanchez, C.A., Dávalos-Rodríguez, I.P., Salazar-Páramo, M., González Lopez, L., Gámez Nava, J.I., and Satoh, M. Autoantibodies to a miRNA-binding protein Argonaute 2 (Su antigen) in patients with hepatitis C virus infection. Clin. Exp. Rheumatol. 2010; 28:842-8.
Pauley, K.M., Satoh, M., Pauley, B.A., Dominguez-Gutierrez, P.R., Wallet, S.M., Holliday, L.S., Cha, S., Reeves, W.H., and Chan, E.K. Formation of GW/P bodies as marker for microRNA-mediated regulation of innate immune signaling in THP-1 cells. Immunol. Cell Biol. 2010; 88:205-12.
Lian, S.L., Li. S., Abadal, G.X., Pauley, B.A., Fritzler, M.J., and Chan, E.K. The C-terminal half of human Ago2 binds to multiple GW-rich regions of GW182 and requires GW182 to mediate silencing. RNA 2009; 15:804-13.
Moser, J.J., Chan, E.K., and Fritzler, M.J. Optimization of immunoprecipitation-western blot analysis in detecting GW182-associated components of GW/P Bodies. Nat Protoc. 2009; 4:674-85.
Nahid, M.A., Pauley, K.M., Satoh, M., and Chan, E.K. miR-146a is critical for endotoxin-induced tolerance: implication on innate immunity. J. Biol. Chem. 2009; 284:34590-34599.
Li, S., Lian, S.L., Moser, J.J., Fritzler, M.L., Fritzler, M.J., Satoh, M., and Chan, E.K. Identification of GW182 and its novel isoform TNGW1 as translational repressors in Ago2 mediated silencing. J. Cell Sci. 2008; 121:4134-44.
Moser, J.J., Eystathioy, T., Chan, E.K., and Fritzler, M.J. Markers of mRNA stabilization, degradation and RNAi within astrocytoma GW bodies. J. Neuroscience Research. 2007; 85:3619-3631.
Bhanji, R.A., Eystathioy, T., Chan, E.K., Bloch, D.B., and Fritzler, M.J. Clinical and serological features of patients with autoantibodies to GW/P Bodies. Clin. Immunol. 2007; 125:247-25.
Lian, S., Fritzler, M.J., Katz, J., Hamazaki, T., Terada, N., Satoh, M., and Chan, E.K. Small interfering RNA-mediated silencing induces target-dependent assembly of GW/P bodies. Mol. Biol. Cell. 2007; 18:3375-87.
Jakymiw, A., Pauley, K.M., Li, S., Lian, S., Ikeda, K., Eystathioy, T., Satoh, M., Fritzler, M.J., and Chan, E.K. The role of GW/P-bodies in RNA processing and silencing. J. Cell Sci. 2007; 120:1317-23.
Ikeda, K., Satoh, M., Pauley, K.M., Fritzler, M.J., Reeves, W.H., and Chan, E.K. Detection of the argonaute protein Ago2 and microRNAs in the RNA induced silencing complex (RISC) using a monoclonal antibody. J. Immunol. Methods. 2006; 317:38-44.
Pauley, K.M., Eystathioy, T., Jakymiw, A., Hamel J.C., Fritzler, M.J., and Chan, E.K. Formation of GW bodies is a consequence of miRNA genesis. EMBO Reports. 2006; 7:904-910.
Jakymiw, A., Ikeda, K., Fritzler, M.J., Reeves, W.H., Satoh, M., and Chan, E.K. Autoimmune targeting of key components of RNA interference. Arthritis Res. Ther. 2006; 8:R87.
Lian, S., Jakymiw, A., Eystathioy, T., Hamel, J.C., Fritzler, M.J., and Chan, E.K. GW bodies, microRNAs and the cell cycle. Cell Cycle. 2006; 5:242-5.
Jakymiw, A., Lian, S., Eystathioy, T., Li, S., Satoh, M., Hamel J.C., Fritzler, M.J., and Chan, E.K. Disruption of GW bodies impairs mammalian RNA Interference. Nature Cell Biology. 2005; 7:1167-74.
Yang, Z., Jakymiw, A., Wood, M.R., Eystathioy, T., Rubin, R.L., Fritzler, M.J., and Chan, E.K. GW182 is critical for the stability of GW bodies expressed during the cell cycle and cell proliferation. J. Cell Science. 2004; 117:5567-78.
Eystathioy, T., Chan, E.K., Mahler, M., Luft, L.M., Fritzler, M.L., and Fritzler, M.J. A panel of monoclonal antibodies to cytoplasmic GW bodies and the mRNA binding protein GW182. Hybridoma & Hybridomics. 2003; 22:79-86.
Eystathioy, T., Jakymiw, A., Chan, E.K., Séraphin, B., Cougot, N., and Fritzler, M.J. The GW182 protein co-localizes with mRNA degradation associated proteins hDcp1 and hLSm4 in cytoplasmic GW bodies. RNA. 2003; 9:1171-1173.
Eystathioy, T., Chan, E.K., Takeuchi, K., Mahler, M., Luft, L.M., Zochodne, D.W., and Fritzler, M.J. Clinical and serological associations of autoantibodies to GW bodies and a novel cytoplasmic autoantigen GW182. J. Mol. Med. 2003; 81:811-818.
Eystathioy, T., Chan, E.K., Tenenbaum, S.A., Keene, J.D., Griffith, K., and Fritzler, M.J. A phosphorylated cytoplasmic autoantigen, GW182, associates with a unique population of human mRNAs within novel cytoplasmic speckles. Mol. Biol. Cell. 2002; 13:1338-51.